

"When light reaches Earth's blue atmospheric cocoon, it encounters its first significant obstacles. Some is reflected back into space or is absorbed by gases, clouds, water vapor, dust, pollen and other ingredients that make up 'air'. But the barriers to free passage of light through the atmosphere are slight compared to the swift changes wrought the moment light enters the sea." – Dr. Sylvia Earle
It's impossible to overstate the importance of well-developed chemosensory capabilities in aquatic species. In the dark, murky, blue-tinted world of the oceans, relying on sight isn't a great sensory strategy, and thus most marine organisms' perceptions of their surroundings are chemical in nature (although sight does play a very important, often quite specialized role in the lives of marine species such as the Caribbean Striped Mantis Shrimp [Lysiosquillina glabriuscula], a "spearer" whose UV vision and fluorescence enable it to catch prey and find mates, and cuttlefish whose ability to see the polarization of light aid in their finely-attuned chromatophoric camouflage). Particularly in invertebrates and protists, signaling molecules and chemical cues mediate nearly every step of reproductive cycles and life history patterns, with chemicals facilitating mate recognition, synchronization in gamete release, and larval settlement. Chemical signals also play a fascinating role in the many commensalistic, parasitic and mutualistic symbioses seen in the ocean; they are of course vital to the mutualistic union that forms the literal and figurative (ecological) backbone for the reef: the coral polyp and its Symbiodinium.

Anthopleura elegantissima
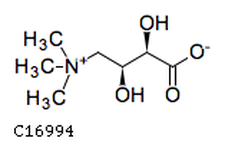 Anthopleuride, an anemone pheromone
Anthopleuride, an anemone pheromone Anthopleura elegantissima serves as a great example of many of these (it is to the study of symbiosis what lab rats are to cancer research or E. coli is to biotechnology), but it's particularly noteworthy for its chemical "warning signals". A. elegantissima is hardly unique in warning conspecifics of predators or alerting them to food sources (a behavior seen in species from swallows to meerkats), nor is it unique in transmitting such as signal chemically (again, seen in many fish and aquatic invertebrates). What's really incredible is how this largely sessile (I say largely sessile because the anemone actually can move along on its base through muscular contractions) cnidarian releases its signaling molecule. The operative pheromone here is anthopleurine, an awesome nitrogen-containing ionic compound (rare is it than an ion serves as the active component of a pheromonal molecule) spectroscopically discovered by Nathan Howe of Stanford's Hopkins Marine Station that triggers responses in anemones at concentrations as low as 35 x 10 to the negative tenth M (as few as 74 picograms/milliliter of water). The anemone's enemy (say that ten time fast!) is the nudibranch Aeolidia papillosa, affectionately dubbed the "shag carpet sea slug" by imaginative malacologists, a species that has harbors fascinating ceratal protozoan symbionts (but we won't talk about those, not yet at least). This miniature (2cm) mollusc munches on A. eleganstissima, in the process taking up large quantities of anthopleurine into its tissue, which subsequently seeps out of the slug as it goes on its way. Anenomes that sense this cloud of anthopleurine immediately convulse protectively, swiftly drawing in their tentacles and closing their mouth. A. papillosa then only has the tough base and column of the anemone to feed on, which has a much higher concentration of anthopleurine, ensuring that the slug won't get another decent meal for a day or two.
Side note: many nudibranchs eat cnidarians, and some aeolid nudibranchs (most notably the species Hermissenda crassicornis) are known to remove the nematocysts from the cnidarian remains in their digestive tract and, through a network of interconnected tubules, deposit them in a "cnidosac" at the tips of the the cerata, where the toxic harpoons can be used to defend against predators. They absorb the strength and toxicity of the cnidarians; they grow stronger on their enemies, the anemones. We still don't know how the nematocysts pass into and through the body of the nudibranch without being discharged.
Side note: many nudibranchs eat cnidarians, and some aeolid nudibranchs (most notably the species Hermissenda crassicornis) are known to remove the nematocysts from the cnidarian remains in their digestive tract and, through a network of interconnected tubules, deposit them in a "cnidosac" at the tips of the the cerata, where the toxic harpoons can be used to defend against predators. They absorb the strength and toxicity of the cnidarians; they grow stronger on their enemies, the anemones. We still don't know how the nematocysts pass into and through the body of the nudibranch without being discharged.

 Navanax inermis
Navanax inermis Another species that exhibits warning signaling is Navanax inermis, a voraciously carnivorous Californian opisthobranch (an older lineage than the nudibranchs). Like A. papillosa (and many other sea slugs, I'm finding), it harbors a symbiont, in this case a parisitic copepod called Anthessius navanacis. These slugs are well known for producing a mucosal slime trail, and when attacked they release from an exocrine gland a yellowish substance into their slime trail containing three pheromones, two of their 3-methyl isomers, and some other minor compounds. At a concentration of 1 x 10 to the negative 5th M, this tips off conspecifics that happen over the yellow-laced slimed trail, and they quickly change course.
What I find really interesting about alarm signaling in N. inermis and A. elegantissima is that neither species is really known for its altruism. Bristle-covered "sensory mounds" on each side of N. inermis's mouth can pick up the trail of almost any undersea critter, including the mucopolysaccharides in the slime trails of conspecifics. If it's hungry enough, N. inermis won't hesitate to follow down a member of its own species and engage in some cannibalism (the slime trails are also a convenient way for males to track down mates). E. elegantissima lives in clonal aggregations, meaning that it's surrounded by asexually-produced clones of itself, and members of the aggregation are friendly enough to each other. But when another aggregation's growth leads to the two groups bumping up against each other, the invaders receive a stinging rebuke. Using their acrorhagi (pale-colored tentacles specialized for repelling rival aggregations), the anemones sting their enemies and leave behind an ectodermic "peel" of skin and stinging nematocysts that kills the invader's tissue.
When it comes finding a mate, pheromones and signaling molecules really are everything to some ocean invertebrates. Mating behavior in many male brachyuran biramous-limbed arthropods (crabs) is initiated by a release of male attractant pheromone. In the case of the Blood-Spotted Swimming Crab, pre-molt females release attractant pheromones through their urine, an NH3 (ammonia) solution produced in the Malphigian tubules. Males spend their time buried in sand and sediment in waters about 10 meters shallower than where the females live, and will not exhibit search behavior unless exposed to the pheromone. Once the male find the female, she stops releasing pheromones, and the male carries her until ecdysis (shedding of the exoskeleton). Males appear to be more aggressive and territorial in colder waters, perhaps explaining why they are kept in the warm, shallow water until needed for egg-fertilizing.
We mentioned the mantis shrimp (stomatopods) above as an example of a marine invertebrate with keen eyesight, but it also has highly developed chemosensory abilities, and signaling chemicals factor into dominance hierarchies associated with mantis shrimp mating. Mantis shrimps are aggressive, and the males engage in ritualized combat over mates. Once one of the males have triumphed, the losers will stay away from the territory he has scented. Mantis shrimp, like many marine crustaceans, capture odorant molecules with chemosensory aesthetsacs on their antennules, which they flick around in the water.
What I find really interesting about alarm signaling in N. inermis and A. elegantissima is that neither species is really known for its altruism. Bristle-covered "sensory mounds" on each side of N. inermis's mouth can pick up the trail of almost any undersea critter, including the mucopolysaccharides in the slime trails of conspecifics. If it's hungry enough, N. inermis won't hesitate to follow down a member of its own species and engage in some cannibalism (the slime trails are also a convenient way for males to track down mates). E. elegantissima lives in clonal aggregations, meaning that it's surrounded by asexually-produced clones of itself, and members of the aggregation are friendly enough to each other. But when another aggregation's growth leads to the two groups bumping up against each other, the invaders receive a stinging rebuke. Using their acrorhagi (pale-colored tentacles specialized for repelling rival aggregations), the anemones sting their enemies and leave behind an ectodermic "peel" of skin and stinging nematocysts that kills the invader's tissue.
When it comes finding a mate, pheromones and signaling molecules really are everything to some ocean invertebrates. Mating behavior in many male brachyuran biramous-limbed arthropods (crabs) is initiated by a release of male attractant pheromone. In the case of the Blood-Spotted Swimming Crab, pre-molt females release attractant pheromones through their urine, an NH3 (ammonia) solution produced in the Malphigian tubules. Males spend their time buried in sand and sediment in waters about 10 meters shallower than where the females live, and will not exhibit search behavior unless exposed to the pheromone. Once the male find the female, she stops releasing pheromones, and the male carries her until ecdysis (shedding of the exoskeleton). Males appear to be more aggressive and territorial in colder waters, perhaps explaining why they are kept in the warm, shallow water until needed for egg-fertilizing.
We mentioned the mantis shrimp (stomatopods) above as an example of a marine invertebrate with keen eyesight, but it also has highly developed chemosensory abilities, and signaling chemicals factor into dominance hierarchies associated with mantis shrimp mating. Mantis shrimps are aggressive, and the males engage in ritualized combat over mates. Once one of the males have triumphed, the losers will stay away from the territory he has scented. Mantis shrimp, like many marine crustaceans, capture odorant molecules with chemosensory aesthetsacs on their antennules, which they flick around in the water.

Two stomatopods fight over prime burrow real estate
 Spongites yendoi, coralline algae
Spongites yendoi, coralline algae Some coral reef species use chemicals to manipulate the behavior of their neighbors in such a way as to increase their chance of survival. One example of such an associational defense is the relationship between non-geniculate crustose coralline algae. Coralline algae have calcareous thalluses, spread out across rocks (or animals' shells) like a splash of paint in the case of non-geniculate species. It has been proven that some species of coralline algae, as well as their bacterial symbionts, produce signaling molecules such as tyrosine and choline derivatives that draw in invertebrate larvae. The larvae will eat epiphytic fleshy algae that compete with the coralline algae for sunlight; in exchange, the coralline algae provides some degree of shelter for the tiny larvae (studies have shown that some mollusc larvae suffer near-100% mortality without the shelter of coralline algae).
Many species exhibit associational chemical defenses, where one species is sheltered by the toxicity of a partner. Chlorodesmis fastigiata is an algae on the Caribbean reef that provides shelter to amphipods, sacoglossans and crabs. These species depend entirely on C. fastigiata for their survival, protected by the algae's chlorodesmin (a highly toxic diterpene). Some carnivorous nudibranchs have also centered their niche around living in the C. fastigia; they feed on the sacoglossans. Tritonia hamnerorum, a Caribbean nudibranch that hangs out around sea fan colonies, sequesters the sea fans' defensive juliannafuran (a type of terpene), rendering the slug unpalatable to fishes.
Many species exhibit associational chemical defenses, where one species is sheltered by the toxicity of a partner. Chlorodesmis fastigiata is an algae on the Caribbean reef that provides shelter to amphipods, sacoglossans and crabs. These species depend entirely on C. fastigiata for their survival, protected by the algae's chlorodesmin (a highly toxic diterpene). Some carnivorous nudibranchs have also centered their niche around living in the C. fastigia; they feed on the sacoglossans. Tritonia hamnerorum, a Caribbean nudibranch that hangs out around sea fan colonies, sequesters the sea fans' defensive juliannafuran (a type of terpene), rendering the slug unpalatable to fishes.
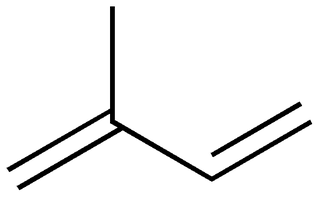
Terpenes are a diverse class of organic compounds, often with strong odors. They are the active component of turpentine (distilled from tree resin, which is full of terpenes), they give hops their aroma and flavor, they act as a cofactor in the biochemical processes behind vision (this cofactor is known as vitamin A, or retinal), and act as building blocks for complex steroids. They are composed of C5H8 (isoprene) units, one of which is depicted in the above figure. Oxidatively modified terpenes are called terpenoids; terpenoids give eucalyptus, cinnamon, cloves and ginger their scent, and the red pigments in tomatoes are a type of terpenoid.
Now it's time to wade into the world of marine epizoic and intracellular symbioses and how these depend on chemical signals. We'll start with epizoic whale barnacles, obligate commensalists formerly grouped under the family Coronulidae (new phylogenetic analysis has revealed some Coronulidae subfamilies to be separate families, including the huge sea-turtle barnacles of the family Chelonibiidae).
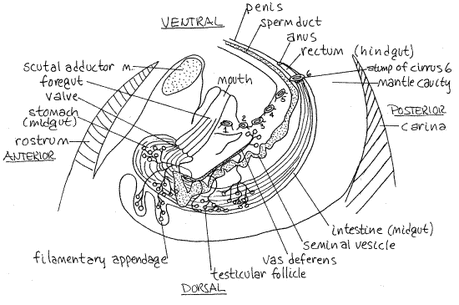
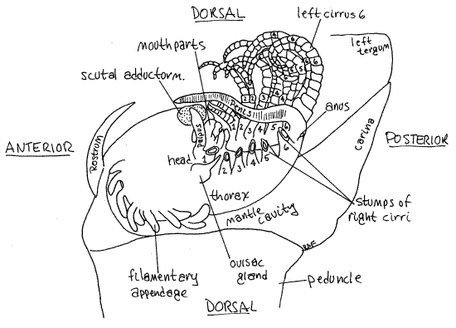
Delightfully detailed renderings of goose barnacle anatomy, from Richard Fox at Lander University. As these figures suggest, barnacles are basically a mouth and a penis attached to a stalk or "peduncle" (inside of which are two ovaries linked to a female gonpore). While barnacles are hermaphroditic, they tend to lean towards one gender and do not generally fertilize themselves; they extend their penises towards a nearby barnacle and deposit sperm in their mantle cavity. According to Chris Neufeld of the Bamfield Marine Sciences Center, "when wave action is light, a longer (thinner) penis can reach more mates, but at times of higher wave action, a shorter (stouter) penis is more manoeuvrable in flow and therefore can reach more mates." Barnacles also mate via spermcasting (the male shoots sperm out into the water and it is taken up by the females).
Barnacles are sessile filter-feeders with calcareous shells, and thus were grouped as molluscs by the otherwise totally awesome Georges Cuvier. The 1830s saw their re-classification as ecdysozoans, and a post-Beagle but pre-Origin of Species Charles Darwin undertook the study and re-classification of hundreds of barnacle species, during which time he coined the term "homology" and cemented his scientific reputation in receiving the Royal Society's Royal Medal for Biology. Up close, barnacles are pretty clearly no molluscs. Their shells are very different, composed of six calcareous plates, they have an exoskeleton, and their embryologic development is very similar to other crustacean arthropods.
Barnacles, sessile creatures with only the simplest of photosensory capabilities, are truly lost without chemical signaling. Their interactions with conspecifics are entirely mediated by chemical cues. A good example is barnacles' release of an a2-macroglobulin-like cuticular protein that draws in cyprids (barnacles in the second nektonic larval stage, after they pass through the five instars of the napulius stage), encouraging the young barnacles to settle close by. For barnacles, living in gregarious (physically clustered) communities is vital to future reproductive success, as these sessile creatures can only mate with neighbors.
Whale barnacles carpet the skin of massive cetaceans, as many as half a ton to a single whale (remember, baleen whales generally weigh in excess of 35 tons; the barnacles are like shorts and a T-shirt to them). The barnacles are referred to as obligate commensalists because 1) the barnacles are highly specialized to whales (in the case of Coronula diadema, Humpbacks specifically) and 2) the relationship is commensal, beneficial to the barnacles but with no net negative loss or gain for the whale. The tiny hangers-on don't harm their massive host in any way, and share in the whale's love of plankton-rich waters (despite the enormous size discrepancy, baleen whales and barnacles are curiously similar from a nutritional perspective: both are filter-feeders with a taste for plankton).
Living on a whale is a highly specialized ecological niche, and it's really very surprising that barnacles ever evolved to live on whales; barnacles were around a hundred million years before cetaceans arose. The whale-hosted lifestyle is confounding for a number of reasons, the first and most obvious being: how do cyprids find a whale in the first place? The answer, as you've probably guessed, is the recognition of chemical cues released from whale skin. In 2006, Yasuyuki Nogata and Kiyotaka Matsumura used a couple of petri dishes and a piece of Humpback skin tissue to prove that cyprid recognition of whales had a chemosensory basis, but we still have no clue what the chemical is that the cyprids are picking up on.
Barnacles, sessile creatures with only the simplest of photosensory capabilities, are truly lost without chemical signaling. Their interactions with conspecifics are entirely mediated by chemical cues. A good example is barnacles' release of an a2-macroglobulin-like cuticular protein that draws in cyprids (barnacles in the second nektonic larval stage, after they pass through the five instars of the napulius stage), encouraging the young barnacles to settle close by. For barnacles, living in gregarious (physically clustered) communities is vital to future reproductive success, as these sessile creatures can only mate with neighbors.
Whale barnacles carpet the skin of massive cetaceans, as many as half a ton to a single whale (remember, baleen whales generally weigh in excess of 35 tons; the barnacles are like shorts and a T-shirt to them). The barnacles are referred to as obligate commensalists because 1) the barnacles are highly specialized to whales (in the case of Coronula diadema, Humpbacks specifically) and 2) the relationship is commensal, beneficial to the barnacles but with no net negative loss or gain for the whale. The tiny hangers-on don't harm their massive host in any way, and share in the whale's love of plankton-rich waters (despite the enormous size discrepancy, baleen whales and barnacles are curiously similar from a nutritional perspective: both are filter-feeders with a taste for plankton).
Living on a whale is a highly specialized ecological niche, and it's really very surprising that barnacles ever evolved to live on whales; barnacles were around a hundred million years before cetaceans arose. The whale-hosted lifestyle is confounding for a number of reasons, the first and most obvious being: how do cyprids find a whale in the first place? The answer, as you've probably guessed, is the recognition of chemical cues released from whale skin. In 2006, Yasuyuki Nogata and Kiyotaka Matsumura used a couple of petri dishes and a piece of Humpback skin tissue to prove that cyprid recognition of whales had a chemosensory basis, but we still have no clue what the chemical is that the cyprids are picking up on.
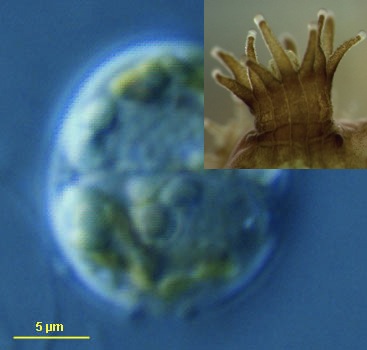
Coral polyps depend on the photosynthates of intracellular algal symbionts for 90% of their sustenance. There are millions of Symbiodinium dinoflagellates in a square centimeter of coral tissue. A Symbiodinium alga has either mastigote (flagellated and motile with an extension of unknown function called the peduncle) or coccoid shape; both types maintain the same organelles. One of the most curious things about the Symbiodinium cell is a reddish-yellow organelle called an accumulation body, possibly used for storing and breaking down wastes.
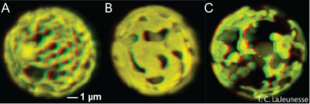
Symbiodinium chloroplasts. Symbiodinium have the same types of chlorophyll as land plants. Symbiodinium also share several of their antenna pigments with land plants, included xanthophylls and carotenes.
The relationship between scleractinian corals and Symbiodinium algae figuratively and literally lays the foundation for the coral reef ecosystem. Coral reefs provide a home to a quarter of marine species, and at the heart of this ecosystem is the coral that feeds, shelters and maintains the biochemical balance of the reef. And the coral, of course, would be nothing without its symbionts – single-celled algae that nourish the coral and give it its coloration. But how is the relationship between host and symbiont established?
Internally brooding corals, which internally fertilize their eggs, generally receive symbionts through maternal inheritance. Once the oocyte is fertilized, it travels to an endodermic pouch and develops into an embryo. The soft mesoglea cloaking the embryo transfers algae from mother to child. But in many corals (including most hermatypic species), fertilization is external, and the planulae (coral larvae) need to find their own symbionts.
Both corals and algae secrete chemicals that function in initial recognition. A study of hydrozoans showed that symbiotic algae that secrete high levels of maltose (the photosynthate polyps feed on) were more likely to be identified and absorbed than algae that could not release maltose. Cnidarians also use pattern-recognitions receptors (PPRs) like lectins to pick up on microbe-associated molecular patterns in algae. C-type lectins are embedded in cellular membranes, and when they bind to passing glycoproteins (like the glycoconjugates released by Symbiodinium) a series of signaling serine proteases are triggered within the coral cell. Coral also secrete lectins (such as the millectin released by Acropora corals and the PdC lectin released by Pocillopora corals) that trigger morphological changes in free-swimming Symbiodinium, preparing them for life as a symbiont. Just like Mycobacterium tuberculosis, Symbiodinium stops the process of phagosomal maturation (being killed as a pathogen and destroyed by lysosomic enzymes) by manipulating endosomal trafficking.
But corals need to keep their algae under control, or the algae will overstay their welcome and become parasites. Corals use signaling peptides termed host-release factors (HRFs) to control the carbon metabolism of their symbionts. HRFs prompt the algae to release glycogen (a form of glucose) and several amino acids. This gives sustenance to the corals while at the same time depriving the algae of nutrients.
(interesting fact: while, like all hydrophilic biological compounds, HRF peptides are very hard to isolate, research indicates that they may be present in many plants and animals and play a role in their carbon metabolism.)
Internally brooding corals, which internally fertilize their eggs, generally receive symbionts through maternal inheritance. Once the oocyte is fertilized, it travels to an endodermic pouch and develops into an embryo. The soft mesoglea cloaking the embryo transfers algae from mother to child. But in many corals (including most hermatypic species), fertilization is external, and the planulae (coral larvae) need to find their own symbionts.
Both corals and algae secrete chemicals that function in initial recognition. A study of hydrozoans showed that symbiotic algae that secrete high levels of maltose (the photosynthate polyps feed on) were more likely to be identified and absorbed than algae that could not release maltose. Cnidarians also use pattern-recognitions receptors (PPRs) like lectins to pick up on microbe-associated molecular patterns in algae. C-type lectins are embedded in cellular membranes, and when they bind to passing glycoproteins (like the glycoconjugates released by Symbiodinium) a series of signaling serine proteases are triggered within the coral cell. Coral also secrete lectins (such as the millectin released by Acropora corals and the PdC lectin released by Pocillopora corals) that trigger morphological changes in free-swimming Symbiodinium, preparing them for life as a symbiont. Just like Mycobacterium tuberculosis, Symbiodinium stops the process of phagosomal maturation (being killed as a pathogen and destroyed by lysosomic enzymes) by manipulating endosomal trafficking.
But corals need to keep their algae under control, or the algae will overstay their welcome and become parasites. Corals use signaling peptides termed host-release factors (HRFs) to control the carbon metabolism of their symbionts. HRFs prompt the algae to release glycogen (a form of glucose) and several amino acids. This gives sustenance to the corals while at the same time depriving the algae of nutrients.
(interesting fact: while, like all hydrophilic biological compounds, HRF peptides are very hard to isolate, research indicates that they may be present in many plants and animals and play a role in their carbon metabolism.)

The fish: Gobidon histrio. The coral: Acropora nasuata, of the abundant Acropora genus of branching corals. The lime-green macro-algae to the goby's left: Chlorodesmis fastigiata, which (as you may remember) packs a poisonous punch in the form of toxic diterpenes. Coral and goby live in a very much chemically-mediated mutualistic partnership: within 15 minutes of contact with the toxic algae, A. nasuata releases chemical cues that bring forth gobies to feast on the toxic bloom. Much like H. crassicornis, the goby takes up toxins from its meal, the algal poison left in its body allowing it to produce toxic secretions that ward off enemies.
Author: Matthew
 RSS Feed
RSS Feed
